¡Descarga Estructura y Funcion de la Célula Humana: Protoplasma, Hidratos de Carbono y Membranas y más Diapositivas en PDF de Fisiología solo en Docsity!
Transporte de sustancias a través de las membranas celulares
El líquido extracelular contiene una gran cantidad de sodio , pero solo una pequeña cantidad de potasio. En el líquido intracelular ocurre lo contrario. Además, el líquido extracelular contiene una gran cantidad de iones cloruro, mientras que el líquido intracelular contiene muy pocos de estos iones. Sin embargo, la concentración de fosfatos y de proteínas del líquido intracelular es considerablemente mayor que la del líquido extracelular. Estas diferencias son muy importantes para la vida de la célula.
Organización de la célula
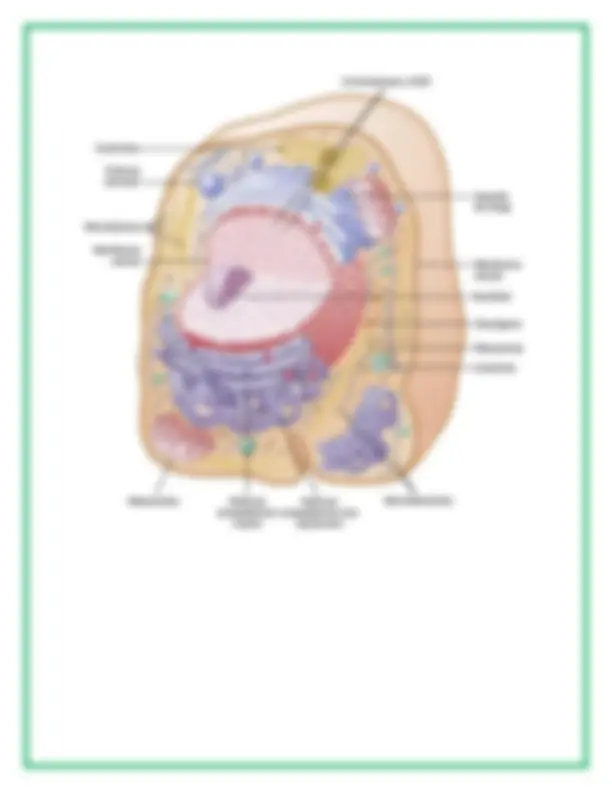
Se muestra una célula típica, tal como se ve en el microscopio óptico. Sus dos partes más importantes son el núcleo y el citoplasma, que están separados entre sí por una membrana nuclear, mientras que el citoplasma está separado de los líquidos circundantes por una membrana celular que también se conoce como membrana plasmática.
Las diferentes sustancias que componen la célula se conocen colectivamente como
protoplasma.
El protoplasma está compuesto principalmente por cinco sustancias: agua,
electrólitos, proteínas, lípidos e hidratos de carbono.
Agua
El principal medio líquido de la célula es el agua, que está presente en la mayoría
de las células, excepto en los adipocitos, en una concentración del 70-85%. Muchos
de los componentes químicos de la célula están disueltos en el agua, mientras que
otros están en suspensión como micropartículas sólidas. Las reacciones químicas
tienen lugar entre los productos químicos disueltos o en las superficies de las
partículas en suspensión o de las membranas.
Iones
Algunos de los iones importantes de la célula son el potasio, el magnesio, el fosfato,
el sulfato, el bicarbonato y cantidades más pequeñas de sodio, cloruro y calcio.
Los iones son los productos químicos inorgánicos de las reacciones celulares y
además son necesarios para el funcionamiento de algunos de los mecanismos de
control celulares.
Por ejemplo, los iones que actúan en la membrana celular son necesarios para la
transmisión de los impulsos electroquímicos en el músculo y las fibras nerviosas.
Proteínas
Después del agua, las sustancias más abundantes en la mayoría de las células son
las proteínas, que normalmente constituyen entre el 10 y el 20% de la masa celular.
Son de dos tipos, proteínas estructurales y proteínas funcionales.
Proteínas estructurales
Están presentes en la célula principalmente en forma de filamentos largos que son
polímeros de muchas moléculas proteicas individuales. Un uso importante de este
tipo de filamentos intracelulares es la formación de microtúbulos que proporcionan
los «citoesqueletos» de orgánulos celulares como los cilios, axones nerviosos,
husos mitóticos de las células en mitosis y masas arremolinadas de túbulos
filamentosos finos que mantienen unidas las partes del citoplasma y nucleoplasma
en sus compartimientos respectivos.
Las proteínas fibrilares se encuentran fuera de la célula, especialmente en las fibras
de colágeno y elastina del tejido conjuntivo y en las paredes de los vasos
sanguíneos, tendones, ligamentos, etc.
No obstante, los hidratos de carbono siempre están presentes en forma de glucosa
disuelta en el líquido extracelular circundante, de forma que es fácilmente accesible
a la célula.
Además, una pequeña cantidad de hidratos de carbono se almacena en las células
en forma de glucógeno, que es un polímero insoluble de glucosa que se puede
despolimerizar y usar rápidamente para aportar la energía que necesitan las células.
Estructura física de la célula
La célula contiene estructuras físicas muy organizadas que se denominan orgánulos intracelulares. La naturaleza física de cada orgánulo es tan importante como lo son los componentes químicos para las funciones de la célula. Por ejemplo, sin uno de los orgánulos, la mitocondria, más del 95% de la energía de la célula que se libera de los nutrientes desaparecería inmediatamente.
La bicapa lipídica básica está formada por tres tipos principales de lípidos: fosfolípidos, esfingolípidos y colesterol.
Los fosfolípidos son los más abundantes en la membrana celular. Un extremo de cada molécula de fosfolípido es soluble en agua, es decir, es hidrófilo, mientras que el otro es soluble solo en grasas, es decir, es hidrófobo. El extremo fosfato del fosfolípido es hidrófilo y la porción del ácido graso es hidrófoba.
Como las porciones hidrófobas de las moléculas de fosfolípidos son repelidas por el agua, pero se atraen mutuamente entre sí, tienen una tendencia natural a unirse unas a otras en la zona media de la membrana.
Las porciones hidrófilas de fosfato constituyen entonces las dos superficies de la membrana celular completa que están en contacto con el agua intracelular en el interior de la membrana y con el agua extracelular en la superficie externa. La capa lipídica de la zona media de la membrana es impermeable a las sustancias hidrosolubles habituales, como iones, glucosa y urea. Por el contrario, las sustancias hidrosolubles, como oxígeno, dióxido de carbono y alcohol, pueden penetrar en esta porción de la membrana con facilidad.
Los esfingolípidos, derivados del aminoalcohol esfingosina, tienen también grupos hidrófobos e hidrófilos y están presentes en pequeñas cantidades en las membranas celulares, especialmente en las células nerviosas. Según se cree, los esfingolípidos complejos de las membranas celulares tienen varias funciones, como son la protección frente a factores perniciosos del entorno, la transmisión de señales y como sitios de adhesión para proteínas extracelulares.
Las moléculas de colesterol de la membrana son también lípidos, porque sus núcleos esteroides son muy liposolubles. Estas moléculas, en cierto sentido, están disueltas en la bicapa de la membrana. Una de sus funciones más importantes consiste en determinar el grado de permeabilidad (o impermeabilidad) de la bicapa ante los componentes hidrosolubles de los líquidos del organismo. El colesterol también controla gran parte de la fluidez de la membrana.
Proteínas de la membrana celular integral y periférica
Estas proteínas de membrana son principalmente glucoproteínas.
Existen dos tipos de proteínas de membrana celular: proteínas integrales que protruyen por toda la membrana y proteínas periféricas que se unen solo a una superficie de la membrana y que no penetran en todo su espesor.
Muchas de las proteínas integrales componen canales estructurales (o poros ) a través de los cuales las moléculas de agua y las sustancias hidrosolubles, especialmente los iones, pueden difundir entre los líquidos extracelular e intracelular. Estos canales de proteínas también tienen propiedades selectivas que permiten la difusión preferente de algunas sustancias con respecto a las demás.
Otras proteínas integrales actúan como proteínas transportadoras de sustancias que, de otro modo, no podrían penetrar en la bicapa lipídica. En ocasiones, estas proteínas transportan incluso sustancias en dirección contraria a sus gradientes electroquímicos de difusión, lo que se conoce como «transporte activo».
Otras proteínas actúan como enzimas. Las proteínas integrales de la membrana pueden actuar también como receptores de los productos químicos hidrosolubles, como las hormonas peptídicas, que no penetran fácilmente en la membrana celular.
Citoplasma y sus orgánulos
El citoplasma está lleno de partículas diminutas y grandes y orgánulos dispersos. La porción de líquido gelatinoso del citoplasma en el que se dispersan las partículas se denomina citosol y contiene principalmente proteínas, electrólitos y glucosa disueltos. En el citoplasma se encuentran dispersos glóbulos de grasa neutra, gránulos de glucógeno, ribosomas, vesículas secretoras y cinco orgánulos especialmente importantes: el retículo endoplásmico, el aparato de Golgi, las mitocondrias, los lisosomas y los peroxisomas.
Citoesqueleto celular: estructuras filamentosas y tubulares
El citoesqueleto celular es una red de proteínas fibrilares organizadas habitualmente en filamentos o túbulos que se originan como moléculas proteicas precursoras sintetizadas por los ribosomas en el citoplasma. Las moléculas precursoras polimerizan después para formar filamentos, por ejemplo, es frecuente que haya grandes cantidades de filamentos de actina en la zona exterior del citoplasma, que se conoce como ectoplasma, para formar un soporte elástico para la membrana celular. Además, los filamentos de actina y miosina se organizan en los miocitos, formando una máquina contráctil especial que es la base de la contracción muscular. Todas las células usan un tipo especial de filamento rígido formado por polímeros de tubulina para construir estructuras tubulares fuertes, los microtúbulos. Una de las funciones principales de los microtúbulos es actuar como citoesqueleto, proporcionando estructuras físicas rígidas para determinadas partes de las células. El citoesqueleto de la célula no solo determina la forma celular sino que además participa en la división de las células, permite su movimiento y proporciona una especie de ruta que dirige el movimiento de los orgánulos en el interior de las células.
Núcleo
El núcleo, que es el centro de control de la célula, envía mensajes a esta para que crezca y madure, se replique o muera. Brevemente, contiene grandes cantidades de ADN, que comprende los genes, que son los que determinan las características de las proteínas celulares, como las proteínas estructurales, y también las enzimas intracelulares que controlan las actividades citoplásmicas y nucleares. Los genes también controlan y promueven la reproducción de la célula. Los genes se reproducen primero para crear dos juegos idénticos de genes y después se divide la célula utilizando un proceso especial, que se conoce como mitosis, para formar dos células hijas, cada una de las cuales recibe uno de los dos juegos de genes de ADN.
Se muestra el aspecto del núcleo en interfase con el microscopio óptico (es decir, en el período entre las mitosis), donde se ve la cromatina, un material que se tiñe de oscuro, por todo el nucleoplasma. Durante la mitosis esta cromatina se organiza en forma de cromosomas muy estructurados.
Membrana nuclear
La membrana nuclear, también conocida como cubierta nuclear, consiste realmente en dos membranas bicapa separadas, una dentro de la otra. La membrana externa es una continuación del retículo endoplásmico del citoplasma celular y el espacio que queda entre las dos membranas nucleares también es una continuación con el espacio del interior del retículo endoplásmico. Varios miles de poros nucleares atraviesan la membrana nuclear. En los bordes de estos poros hay unidos grandes complejos de moléculas proteicas, de forma que la zona central de cada poro mide solo unos 9 nm de diámetro, tamaño suficientemente grande como para permitir que moléculas de un peso molecular de hasta 44.000 la atraviesen con una facilidad razonable.
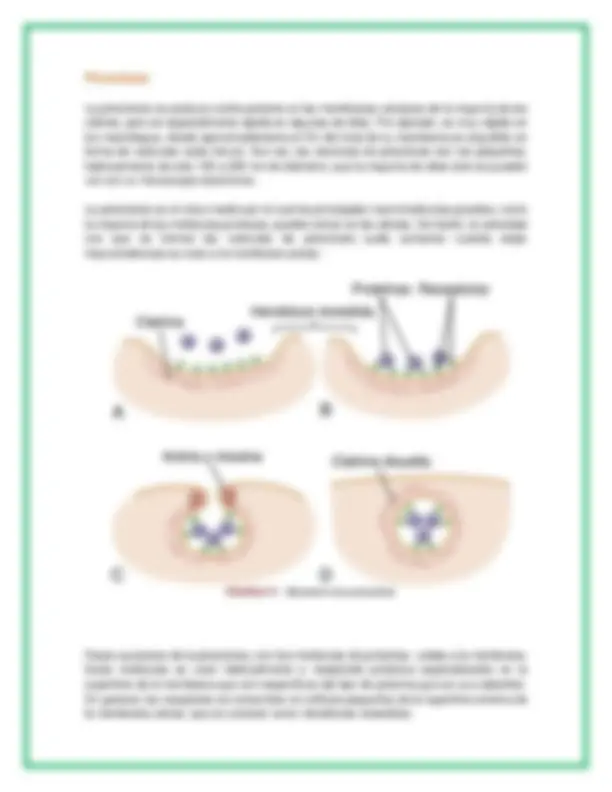
Pinocitosis
La pinocitosis se produce continuamente en las membranas celulares de la mayoría de las células, pero es especialmente rápida en algunas de ellas. Por ejemplo, es muy rápida en los macrófagos, donde aproximadamente el 3% del total de su membrana es engullido en forma de vesículas cada minuto. Aun así, las vesículas de pinocitosis son tan pequeñas, habitualmente de solo 100 a 200 nm de diámetro, que la mayoría de ellas solo se pueden ver con un microscopio electrónico.
La pinocitosis es el único medio por el cual las principales macromoléculas grandes, como la mayoría de las moléculas proteicas, pueden entrar en las células. De hecho, la velocidad con que se forman las vesículas de pinocitosis suele aumentar cuando estas macromoléculas se unen a la membrana celular.
Pasos sucesivos de la pinocitosis, con tres moléculas de proteínas unidas a la membrana. Estas moléculas se unen habitualmente a receptores proteicos especializados en la superficie de la membrana que son específicos del tipo de proteína que se va a absorber. En general, los receptores se concentran en orificios pequeños de la superficie externa de la membrana celular, que se conocen como hendiduras revestidas.
En el interior de la membrana celular, por debajo de estas hendiduras, hay una red de una proteína fibrilar conocida como clatrina, así como otras proteínas, quizás incluso filamentos contráctiles de actina y miosina.
Una vez que las moléculas proteicas se han unido a los receptores, las propiedades de superficie de esa zona de la membrana cambian de tal forma que todas las hendiduras se invaginan hacia el interior y las proteínas fibrilares que rodean a la que se invagina hacen que se cierren los bordes sobre las proteínas unidas y sobre una pequeña cantidad de líquido extracelular. Inmediatamente después la porción invaginada de la membrana se rompe separándose de la superficie de la célula, formando una vesícula de pinocitosis dentro del citoplasma de la célula.
Lo que hace que la membrana celular realice las contorsiones necesarias para formar las vesículas de pinocitosis sigue sin estar claro. Este proceso requiere el aporte de energía desde el interior de la célula, que es suministrada por el ATP.
Este proceso requiere, además, la presencia del ion calcio en el líquido extracelular, que probablemente reaccionará con los filamentos de proteína contráctil que hay por debajo de las hendiduras revestidas para proporcionar la fuerza que se necesita para que se produzca la separación de las vesículas lejos de la membrana celular.
Fagocitosis La fagocitosis se produce, a grandes rasgos, del mismo modo que la pinocitosis, excepto porque implica la participación de partículas grandes y no moléculas. Solo algunas células tienen la capacidad de realizar la fagocitosis, principalmente los macrófagos tisulares y algunos leucocitos sanguíneos. La fagocitosis se inicia cuando una partícula, como una bacteria, una célula muerta o un resto de tejido, se une a los receptores de la superficie de los fagocitos. En el caso de las bacterias, cada una de ellas ya suele estar unida a un anticuerpo específico frente a ese organismo y es ese anticuerpo el que se une a los receptores de fagocitosis, arrastrando consigo a la bacteria. Esta intermediación de los anticuerpos se conoce como opsonización,
La fagocitosis se produce en las etapas siguientes:
- Los receptores de la membrana celular se unen a los ligandos de superficie de la partícula.
- La zona de la membrana alrededor de los puntos de unión se invagina hacia fuera en una fracción de segundo para rodear a toda la partícula, y después cada vez más receptores de membrana se unen a los ligandos de la partícula. Todo esto ocurre bruscamente, como si fuera una cremallera, para formar una vesícula fagocítica cerrada.
- La actina y otras fibrillas contráctiles del citoplasma rodean la vesícula fagocítica y se contraen en torno a su borde exterior, empujando la vesícula hacia el interior.
aminoácidos, glucosa, fosfatos, etc., que pueden difundir a través de la membrana de las vesículas hacia el citoplasma. Lo que queda en la vesícula digestiva, que se denomina cuerpo residual, representa las sustancias indigestibles. En la mayoría de los casos, el cuerpo residual se excreta finamente a través de la membrana celular en un proceso que se denomina exocitosis, que es esencialmente lo contrario que la endocitosis. Es decir, las vesículas de pinocitosis y fagocitosis que contienen los lisosomas pueden considerarse los órganos digestivos de las células.
Regresión de los tejidos y autólisis de las células dañadas
Los tejidos del organismo a menudo regresan a un tamaño más pequeño. Esta regresión se da, por ejemplo, en el útero después del embarazo, en los músculos tras períodos prolongados de inactividad y en las glándulas mamarias al final de la lactancia. Los lisosomas son responsables de gran parte de esta regresión. Otro papel especial de los lisosomas es la eliminación de las células o porciones de células dañadas en los tejidos. El daño de una célula causado por el calor, el frío, un traumatismo, productos químicos o cualquier otro factor induce la rotura de los lisosomas. Las hidrolasas liberadas comienzan inmediatamente a digerir las sustancias orgánicas circundantes. Si el daño es pequeño, solo se eliminará una porción de la célula, que después se repararía. Si el daño es importante se digiere toda la célula, lo que se denomina autólisis. De esta manera, la célula se elimina por completo y se forma una célula nueva del mismo tipo, normalmente por la reproducción mitótica de una célula adyacente para ocupar el puesto de la anterior.
Los lisosomas también contienen sustancias bactericidas que pueden matar a las bacterias fagocitadas antes de que provoquen daños a la célula. Estas sustancias son:
- la lisozima, que disuelve la membrana celular bacteriana;
- la lisoferrina, que se une al hierro y a otras sustancias antes de que puedan promover el crecimiento bacteriano, y
- un medio ácido, con un pH en torno a 5, que activa las hidrolasas e inactiva los sistemas metabólicos bacterianos.
Reciclado de los orgánulos celulares: autofagia
Los lisosomas desempeñan un papel fundamental en el proceso de autofagia, que literalmente significa «comerse a sí mismo». La autofagia es un proceso de limpieza según el cual los orgánulos y los grandes agregados proteicos obsoletos se degradan y se reciclan